












【佳学基因检测】纤毛的结构及超微结构:研究生入学必考知识点
纤毛的基本结构
纤毛的超微结构在整个进化过程中令人谅呀地得到了惊人的保存。 初级纤毛和运动纤毛的核心(轴丝)由位于纤毛外围的九个微管双联体组成; 这些微管与基体的九个微管三联体中的三个微管中的两个连续,基体是一种将纤毛固定在细胞体上并通过过渡纤维附着在细胞膜上的结构。 活动纤毛(以及由特定细胞类型组装的不动纤毛)的核心还包含两个位于中央的单个微管:C1 和 C2(9 + 2 配置,图 1)。 活动纤毛也有许多附着在外周和中央微管上的多蛋白复合物。 中央微管及其复合体,即所谓的投射(C1a-C1f 和 C2a-C2e),以及连接两个微管的复合体(桥)形成一个中央装置。 据信,调节纤毛跳动的部分信号起源于这个中央结构。
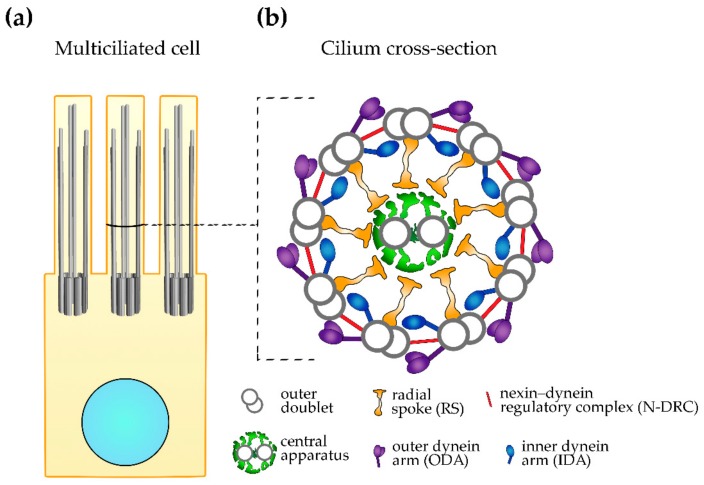
图1:运动纤毛超微结构。 (a) 具有标记核(浅蓝色)、基体和纤毛的多纤毛细胞示意图。 (b) 纤毛横截面示意图,显示大的纤毛复合体((a) 中标记的横截面水平)。
外双微管是四种大型复合体的停靠位点:外动力蛋白臂和内动力蛋白臂(分别为 ODA 和 IDA)、连接蛋白-动力蛋白调节复合物 (N-DRC) 和径向辐条 (RSs),以及许多主要由 未知的蛋白质组成和功能 [12,15,16]。 这些大复合体以及小纤毛复合体沿外周微管排列,形成由 96 nm 单位组成的特征模式,称为轴丝重复; 每个重复序列有四个相同的 ODA(双头或三头,取决于生物体)、七个 IDA(一个双头和六个单头,都包含不同的动力蛋白)、一个 N-DRC、三个略有不同的 RS 它们的结构和可能的功能,以及小型复合体的单个副本(例如,tether/tetherhead、MIA 复合体)[15,16,17,18,19]。
径向辐条在中央装置的方向上延伸。 有人提出,径向辐条和中央装置的投影之间的瞬时接触有助于将源自中央装置的信号传输到外部双臂和动力蛋白臂,从而调节纤毛跳动 [20]。
外动力蛋白臂和内动力蛋白臂由许多蛋白质亚基组成,包括动力蛋白臂类型特异性负端定向马达、动力蛋白。 激活的动力蛋白沿相邻微管的时空同步运动导致相邻双峰的移动 [21]。 这种转变的范围受到 N-DRC 的限制,它在两个相邻的双峰之间延伸。 因此,外部双峰的移动被转化为纤毛弯曲,随后弯曲为纤毛跳动。
可以使用经典的透射电子显微镜 (TEM) 观察大型外部双峰复合体,但小型复合体或大型复合体结构的微小变化低于 TEM 检测水平。
真核鞭毛中央对复合体中的保守结构基序
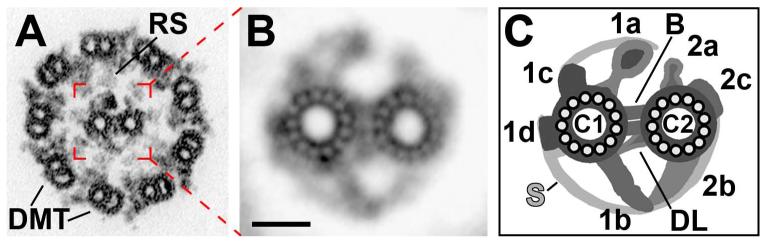
人体结构与功能的基因解码揭示活动纤毛和鞭毛是高度保守的细胞器,在真核细胞中产生动力、感知环境和发出信号。 纤毛和鞭毛的缺陷是人类多种疾病的产生原因,这一类疾病在《人体基因序列变化与疾病表征》中被称为,称为纤毛病。 几乎所有活动的纤毛和鞭毛都具有由微管 (MT) 形成的核心结构,即轴丝。它是由 9+2 排列的 9 个双联系微管 (DMT) 围绕一对称为中央对复合体 (CPC) 的单体微管组成。 为了产生纤毛和鞭毛的特征性的波形运动,400 多个轴丝组件的正确组装并发挥正常的和功能是必要,单个蛋白质的突变可导致纤毛/鞭毛瘫痪,例如 PF16。在结构上,每个双联体微管( DMT) 都是由一个 96 nm 长的结构单元不断累积形成的,该结构单体沿 DMT 纵轴重复。 这个 96 nm 重复单元中的关键组件是动力蛋白马达,它们在双联体微管上的 A 管上排列成两排,即内动力蛋白臂和外动力蛋白臂(IDA 和 ODA)。 这些动力蛋白有效附着锚定在一个双联系微管上上,并利用 ATP 的能量通过沿着相邻的双联系微管行走产生运动所需的机械力,从而导致双联体微管相互滑过。 这种双联体微管间的滑动是有限的,例如 通过连接相邻双联体微管的连接蛋白-动力蛋白调节复合物 (N-DRC) 的连接蛋白链接,将双联体微管之间的线性滑动转化为纤毛弯曲。 为了产生典型的纤毛和鞭毛的击打模式,在任何给定时间只有部分动力蛋白必须是活跃的; 同时激活所有动力蛋白会使轴丝僵硬且无法运动。 动力蛋白的协调是通过整合来自轴丝调节复合体的各种信号来实现的,例如径向辐条 (RS)、I1 内部动力蛋白、N-DRC 和 CPC。
不同生物的纤毛和鞭毛已被用于研究 CPC,包括来自原生生物(例如衣藻和草履虫)、棘皮动物(例如 Strongylocentrotus 精子)和哺乳动物(例如小鼠精子和上皮细胞以及人类精子鞭毛)的纤毛和鞭毛。 通常,CPC 似乎具有保守的基本结构,如图 1 所示:两个单态 MT,称为 C1 和 C2,通过统称为桥的结构连接在一起。 附加到每个 MT 的是向 RS 和 DMT 延伸的投影。 对 CPC 结构产生不利影响的突变范围从一个投影的丢失到 CPC 组装的有效失败。 单个投射的丢失会导致鞭毛跳动率显着降低,而一个 MT 或整个 CPC 的缺失会导致更严重的鞭毛麻痹表型,这表明 CPC 是运动的重要调节器。 CPC 作为运动调节剂的这种作用似乎是保守的,因为同源 CPC 基因的突变与各种以不动鞭毛/纤毛为特征的人类纤毛病有关,例如原发性纤毛运动障碍、不孕症和脑积水。
径向丝的结构与功能
大多数活动的纤毛和鞭毛由 9 个含动力蛋白臂的外周双微管 (DMT) 组成,围绕着一对中央微管 (CP)(“9+2”轴丝)。 径向辐条 (RS) 是一种 T 形蛋白质复合物,具有指向 CP 的正交头部和锚定在 DMT 的每个 A 管上的茎。 它充当 CP 和轴丝动力蛋白臂之间的机械化学传感器,以调节鞭毛/纤毛运动。 广泛使用的模式生物莱茵衣藻的鞭毛在轴丝的每个 96 nm 重复单元中包含两个全尺寸 RS(RS1 和 RS2)。 相比之下,嗜热四膜虫和后生动物的活动纤毛/鞭毛具有三联体 RS(RS1 至 RS3)。 衣藻 RS 由至少 23 个亚基蛋白(RSP1 至 RSP23)组成。 其中 17 个具有哺乳动物同系物。 导致整个 RS 或 RS 头丢失的突变导致衣藻的鞭毛不动,但在哺乳动物中导致纤毛旋转,导致原发性纤毛运动障碍 (PCD),这是一种以反复呼吸道感染、内脏反位、不育和脑积水为特征的遗传综合征 .
RS 中贼显着的形态差异在于 RS 头部,这是通过直接接触 CP 的投影来调节机械信号的关键结构域。 RS1 和 RS2 的头部由两个结构相同、旋转对称的两半组成,与 RS3 的头部有很大不同。 此外,它们的形态在原生动物和后生动物之间显着不同。 例如,在衣藻和四膜虫中,RS1 和 RS2 的头部富含侧枝,这些侧枝也在两个头部之间形成连接。 相比之下,在海胆 (Strongylocentrotus purpuratus) 和人类中,RS1 和 RS2 的头部类似于一对溜冰刀,朝向 CP 的接口少得多。 尽管 RS 和 RS 头在纤毛/鞭毛运动中很重要,但 RS 和 RS-CP 相互作用的结构细节仍然知之甚少,尤其是在哺乳动物中。
RS 头可能已经过改造以符合进化过程中轴丝的结构和功能变化。 然而,形态变化是如何发生的仍不清楚。 衣藻RS头由RSP1、-4、-6、-9、-10和茎部RSP3的一部分(C端)组成。 头部的每一半对称部分都包含这些组件的一个副本。 所有的头部成分都具有哺乳动物直系同源物(Rsph1、-4a、-6a、-9、-10b 和 -3b)。 与后生动物 RS 头显着减少的表面积形成鲜明对比的是,人类 RSPH4A、-6A 和 -10B 的肽分别比其衣藻同源物长 1.5、1.3 和 4 倍。 只有 RSPH1(309 个氨基酸 [aa])比 RSP1(814 个氨基酸)短 (11)。 小鼠 RS 头蛋白的长度也与其人类对应物类似地发生了变化。 此外,虽然小鼠 Rsph4a 对于气管、室管膜和输卵管的活动多纤毛中 RS1 至 RS3 的头部形成至关重要,但 Rsph6a 在精子中的正常鞭毛形成中特异性表达。 RSP4/Rsph4a 和 RSP6/Rsph6a 是旁系同源物:RSP4 和 RSP6 具有 48% 的序列同一性,而鼠 Rsph4a 与 Rsph6a 有 63% 的同一性。 然而,海胆和玻璃海鞘只有一个直系同源物。 这些结果表明,与原生动物 RS 头部不同,后生动物头部可能不会同时包含 Rsph4a 和 Rsph6a。 轴丝中 RS 结构的一般形状已通过传统电子显微镜 (EM) 和低温电子断层扫描 (cryo-ET) 确定。 贼近,通过冷冻电镜单粒子分析解决了衣藻的 15 Å 分辨率 RS 结构。 然而,分辨率不足以描述单个 RS 亚基的位置。
佳学基因基因纤毛病的结构与功能基因解码,通过生化和结构分析,表明鼠类 RS 头部在成分和形态上都不同于衣藻。 基因解码研究表明,RS 头部经历了深刻的重塑,可能在协调纤毛或鞭毛运动的进化过程中符合轴丝的结构和功能改变。
- 来了,就说两句!
-
- 贼新评论 进入详细评论页>>
