












【佳学基因检测】在儿科检测两种 HLA II 基因变异预测骨肉瘤风险
HLA突变与骨肉瘤风险基因检测导读:
背景:
尽管发表了全基因组关联研究,但骨肉瘤的遗传病因仍然知之甚少。已经观察到人类白细胞抗原 (HLA) 基因变异与几种癌症风险之间的关联,但标准 SNP 阵列无法很好地捕获 HLA 变异。
方法:
骨肉瘤风险基因检测方案开发小组使用定制的全基因组阵列对 207 名加利福尼亚小儿骨肉瘤病例和 696 名欧洲血统对照进行了基因分型,并在 MHC 区域内补充了约 6,000 个额外的探针。随后,骨肉瘤风险基因检测方案开发小组使用 5,225 名通过下一代测序进行高分辨率 HLA 分型的参考小组来估算四位数的经典 HLA 等位基因。病例对照比较针对具有祖先信息的主成分进行了调整,发现分析中的顶级关联在 657 例病例和 1183 例对照的独立数据集中进行了复制。
结果:
在发现分析中,三个高度相关的 HLA II 类变异 (r 2 =0.33–0.98) 与骨肉瘤风险相关,包括 HLA-DRB1*0301 (OR=0.52; P=3.2×10 -3 )、HLA-DQA1*0501 ( OR=0.74;P=0.031)和HLA-DQB1*0201(OR=0.51;P=2.7×10 -3)。在复制数据中观察到类似的关联(P范围= 0.011-0.037)。两个数据集的荟萃分析将 HLA-DRB1*0301 确定为最显着相关的变体(OR meta =0.62;P meta =1.5×10 -4),达到 Bonferroni 校正的统计显着性。荟萃分析还揭示了 HLA-DQA1*01:01 处的第二个重要独立信号(OR meta =1.33,Pmeta =1.2×10 -3),以及在 HLA-DQB1*0302 处的第三个暗示关联(OR meta =0.73, P meta =6.4×10 -3)。
结论:
多个独立的 HLA II 类等位基因可能会影响骨肉瘤的风险。
影响:
需要做更多的工作来将骨肉瘤风险基因检测方案开发小组的观察扩展到其他患者群体,并阐明这些关联背后的潜在因果机制。了解免疫对骨肉瘤病因的贡献可能会为合理的治疗靶点提供信息。
介绍
骨肉瘤是最常诊断的原发性恶性骨肿瘤,发病高峰发生在青春期。在美国,每年在 20 岁以下的个体中诊断出大约 400 例新病例 ,在全球范围内观察到的儿童期和青少年期骨肉瘤发病率相似 。骨肉瘤的危险因素包括较高的身材和男性,但该疾病的遗传病因仍然知之甚少。虽然骨肉瘤风险增加与骨佩吉特病和遗传性癌症易感综合征(例如Li-Fraumeni 综合征)有关,但大多数病例是散发性的. 候选基因研究表明,DNA 修复途径 、生长激素途径 和端粒维持途径 中的常见遗传变异在导致骨肉瘤风险中的作用,但只有一个单一的全基因组关联研究 (GWAS) 迄今已发表 。该 GWAS 报告了两个与骨肉瘤相关的风险位点,其中一个位于GRM4基因的 6p21.3 处,距主要组织相容性复合体 (MHC) II 类区域约 1 Mb。
MHC 区域,在染色体 6p21 上包含约 7.6Mb,是基因组中多态性和基因最密集的区域之一,包括高度可变的人类白细胞抗原 (HLA) 基因。在之前对慢性淋巴细胞白血病、肺鳞状细胞癌、卵巢癌、乳腺癌、宫颈癌和鼻咽癌的研究中,HLA 变异与癌症风险相关。已经假设了将 HLA 遗传变异与癌症发展联系起来的各种机制,包括免疫系统对肿瘤的不同耐受性、免疫监视的效率和癌症免疫编辑导致某些 HLA 变异易患特定恶性肿瘤。研究可遗传的 HLA 变异在骨肉瘤风险中的作用可能会为疾病病因学提供信息,并为免疫治疗的潜在策略提供信息。
虽然之前的骨肉瘤 GWAS 没有报告 MHC 区域的任何信号,但由于 MHC 中序列和拷贝数变异的独特特征,标准全基因组 SNP 阵列不能很好地捕获 HLA 基因中的遗传变异和单倍型结构地区。使用定制的 Affymetrix Axiom 阵列,并辅以跨越 MHC 区域的约 6,000 个额外探针,骨肉瘤风险基因检测方案开发小组以四位数分辨率键入经典 HLA 等位基因(例如 HLA-DRB1*1501) 并测试与小儿骨肉瘤风险的关联。骨肉瘤风险基因检测方案开发小组随后尝试在一组独立的骨肉瘤病例和对照中进行复制,并对这两个数据集进行荟萃分析,总共 864 例病例和 1879 例对照。骨肉瘤风险基因检测方案开发小组进一步评估了 MHC 区域中的非加性遗传关联作为一种补充方法,因为观察到 HLA 变体对许多疾病的广泛非加性和上位效应,以及受试者性别的任何潜在修饰或临床表现(例如肿瘤位置、转移的存在、诊断时的年龄)以进一步了解这种侵袭性癌症的病因、进展和预后。
材料和方法
发现样本:
该研究得到了加州大学伯克利分校和旧金山分校的机构审查委员会以及加州公共卫生部 (CDPH) 的批准。本研究中使用的生物样本和/或数据来自加利福尼亚生物银行计划(SIS 请求编号 550)。CDPH,遗传疾病筛查部门从该州出生的所有新生儿中获取新生儿血液样本,以进行疾病筛查。自 1982 年以来,筛选后残留的血斑已在 -20°C 下存档,并可供批准的研究使用。骨肉瘤风险基因检测方案开发小组将 CDPH 维护的全州出生记录(1982-2009 年)与加州癌症登记处(CCR,1988-2011 年)的癌症诊断数据联系起来。根据 CCR 记录,该分析包括 1982 年至 2009 年期间在加利福尼亚出生并在 19 岁时被诊断出患有骨肉瘤(ICD-O-3 代码 9180-9183、9185-9187 和 9192-9195)的非西班牙裔白人儿童。选择同一时期在加利福尼亚州出生且未向 CCR 报告患有任何儿童癌症的儿童作为对照。
DNA提取:
将 12mm 干血斑的三分之一分成三个均匀的部分,并在提取前置于 2mL 微量离心管中,使用 QIAamp DNA Investigator Kit (Qiagen) 进行提取。简而言之,将 280 μL Buffer ATL 和 20 μL 蛋白酶 K 添加到每个样品中。涡旋样品,然后在干浴振荡器中以 900rpm 和 56°C 孵育一小时。孵育后,将样品短暂离心,将裂解液转移到新的 2mL 微量离心管中,同时丢弃固体残留物。将 1μL 的 1ng/μL 载体 RNA 添加到裂解物中,然后短暂涡旋。加入载体 RNA 后,将样品放入 Qiagen Qiacube 自动化工作站进行 DNA 分离。Qiacube 提取方案的结果是 ATE 缓冲液中的纯化 DNA 样品。
Discovery 样本基因分型和质量控制:
根据病例对照状态、报告的种族和性别,使用封闭式随机化将 DNA 标本分配到基因分型板。DNA 在 Affymetrix Axiom Latino Array 上进行基因分型,具有高覆盖率的欧洲血统,并在 MHC 区域补充了约 6,000 个额外的 SNP 探针。DNA 样本在 Affymetrix TITAN 系统上进行基因分型,并使用 Affymetrix Genetools 处理原始图像文件以调用基因型。
阵列中包含的重复样本 (n = 34) 的平均基因型一致性 > 99%。如前所述 迭代地执行 SNP 和样本的调用率过滤,不包括调用率 <97% 的 SNP 和调用率 <97% 的样本。在欧洲血统对照和报告性别与基因分型性别不匹配的样本中,SNP显示出明显偏离Hardy-Weinberg平衡(P<1.0×10 -5 )被排除在外。发现数据中排除了来自每个受试者对的一个个体,其身份认同(IBD)比例> 0.18。使用来自 1184 个 HapMap Phase 3 样本的全基因组 SNP 阵列数据,骨肉瘤风险基因检测方案开发小组使用等位基因频率>0.05 的未链接常染色体双等位基因 SNP 进行主成分分析,并从分析中删除任何显示非欧洲血统证据的样本(平均 CEPH 值 >3 SD在 PC 1-3 上)。
复制数据集:
骨肉瘤复制数据集来自 dbGaP 研究加入 phs000734.v1.p1(骨肉瘤风险的全基因组关联研究 (GWAS))和 phs000381.v1.p1(eMERGE Geisinger eGenomic Medicine MyCode Project Controls)。病例和对照在 Illumina OmniExpress 阵列上进行基因分型,并如前所述进行质量控制过滤 。简而言之,调用率 <0.98 的 SNP 和基因分型调用率 <0.97 的受试者被删除。使用 Eigenstrat 和 HapMap 参考样本和前五个主成分的平均值是在 HapMap CEPH 样本中计算的。具有非欧洲血统证据的受试者(平均 CEPH 值 > 3 SD)被排除在外。具有 Hardy-Weinberg 平衡P <1.0×10 -5的 SNP其中控件被删除。比较了发现和复制数据集中的骨肉瘤病例,并从复制数据集中排除了重复和隐匿相关(IBD>0.18)样本(n=22 重复患者,0 例隐匿相关患者)。最终复制数据集中总共包含 657 个不重叠的欧洲血统病例和 1183 个对照。来自 dbGaP 的骨肉瘤患者主要是儿童和青少年(年龄 < 21 岁),尽管无法获得个体水平的年龄数据,并且最初的出版物并不限于特定的年龄范围。这些骨肉瘤患者是先前 GWAS 出版物中包含的患者的一个子集 。
HLA基因型的插补:
骨肉瘤风险基因检测方案开发小组估算了三个主要 I 类基因(HLA-A、HLA-B 和 HLA-C)和五个 II 类基因(HLA-DRB1、-DPA1、-DPB1、-DQA1 和-DQB1),以及使用 SNP2HLA 的 5,695 个 HLA 基因内 SNP,它使用来自 1 型糖尿病遗传学联盟的 5,225 名欧洲血统个体的参考小组,这些人通过下一代测序进行了高分辨率 HLA 分型。如前所述,骨肉瘤风险基因检测方案开发小组使用 SNP2HLA 推算的等位基因剂量数据进行主要的 MHC 范围关联分析,并使用分阶段的最佳猜测基因型进行非加性效应分析。关联分析仅限于经典的四位数 HLA I 类和 II 类等位基因,插补质量评分 INFO>0.80,等位基因频率>0.05(19 个 I 类等位基因,27 个 II 类等位基因)。对于非加性效应分析,骨肉瘤风险基因检测方案开发小组仅限于在每个分析的 HLA 基因上具有两个最佳猜测单倍型的个体。
统计分析:
骨肉瘤风险基因检测方案开发小组对 HLA I 类和 II 类等位基因中的每一个进行了逻辑回归分析,使用 SNP2HLA 输出的推算等位基因剂量和 PLINK 1.9 中的“逻辑”命令,调整 Eigenstrat 生成的前 10 个祖先信息主成分。作为二次探索性调查,骨肉瘤风险基因检测方案开发小组还对 MHC 区域中的 5,695 个估算的 HLA 基因内 SNP 进行了关联分析。分析在发现和复制数据集中分别进行,并使用 PLINK 中的“元分析”命令进行元分析。骨肉瘤风险基因检测方案开发小组通过重复调整顶部信号的逻辑回归来进行条件分析。骨肉瘤风险基因检测方案开发小组还针对 SNP rs1906953 进行了调整,这是一种先前发现的骨肉瘤 GWAS 在 6p21 命中。3 位于距 MHC 区域约 1 Mb 的位置,以评估骨肉瘤风险基因检测方案开发小组分析中存在的顶部信号的任何衰减。对于性别分层分析,骨肉瘤风险基因检测方案开发小组分别检查了男性和女性的关联信号,并测试了效应的异质性。骨肉瘤风险基因检测方案开发小组还进行了仅病例分析,评估病例对照分析中确定的顶级 HLA 关联信号是否与临床表现的差异相关,包括:诊断年龄、肿瘤位置、肿瘤大小、扩展、分化和转移的存在。有关临床变量的信息仅可用于发现数据集,编码如前所述(评估病例对照分析中确定的顶级 HLA 关联信号是否与临床表现的差异相关,包括:诊断年龄、肿瘤位置、肿瘤大小、扩展、分化和转移的存在。有关临床变量的信息仅可用于发现数据集,编码如前所述(评估病例对照分析中确定的顶级 HLA 关联信号是否与临床表现的差异相关,包括:诊断年龄、肿瘤位置、肿瘤大小、扩展、分化和转移的存在。有关临床变量的信息仅可用于发现数据集,编码如前所述。
骨肉瘤风险基因检测方案开发小组通过评估先前研究中描述的优势效应模型来研究 HLA 等位基因的非加性效应 。简而言之,骨肉瘤风险基因检测方案开发小组测试了模型拟合的改进,将加性效应模型与另外包括一个表示所研究的每个 HLA 等位基因的杂合状态的显性项的模型进行比较。来自 χ2 检验的模型改进的 P 值表示与加性模型的偏差的显着性。骨肉瘤风险基因检测方案开发小组还测试了检测到的每个独立信号中的顶级变体与每个剩余的 II 类变体之间的相互作用。
在发现分析中达到标称统计学意义的 HLA I 类和 II 类等位基因 (P<0.05) 被用于复制。在两个数据集中实现名义显着性并且在荟萃分析中实现超过 Bonferroni 校正的 P 值的变体被认为具有统计学意义。由于 HLA 变体之间的连锁不平衡 (LD),传统的 Bonferroni 校正阈值将过于保守,因为测试不是独立的。骨肉瘤风险基因检测方案开发小组估计了在调整 LD 以校正多重比较后进行的独立测试,将“有效”测试的数量从 46 个主要 HLA I 类和 II 类等位基因分析减少到 39 个,将次要 HLA 基因内 SNP 分析从 5,695 个减少到 1,713 个。然后使用 HLA I 类和 II 类等位基因(0.05/39=1.3×10 -3)和 HLA 基因内 SNP(0.05/1713=2.9×10 -5)。
结果
发现数据集中共有 13,103 个 MHC 区域的 SNP 在阵列上成功进行基因分型,其中包括 207 例小儿骨肉瘤病例和 696 例欧洲血统对照。使用这些数据来估算 HLA I 类和 II 类等位基因,骨肉瘤风险基因检测方案开发小组观察到 19 个 I 类等位基因和 27 个 II 类等位基因,频率>0.05。在逻辑回归分析中,三个 I 类等位基因名义上与骨肉瘤风险相关,包括:HLA-A*0201(OR=1.35;95% CI=1.05-1.74;P=0.020)、HLA-A*0301(OR=0.66) ;95% CI=0.46–0.94;P=0.023)和 HLA-B*0801(OR=0.60;95% CI=0.38–0.92;P=0.021)(表1)。此外,三个 II 类等位基因名义上与骨肉瘤风险相关,包括 HLA-DRB1*0301 (OR=0.52;95% CI=0.34–0.81;P=3.2×10 -3 )、HLA-DQA1*0501 (OR=0.74 ;95% CI=0.56–0.97;P=0.031)和HLA-DQB1*0201(OR=0.51;95% CI=0.33–0.79;P=2.7×10 -3)(表1)。
表1:207例骨肉瘤病例和696例对照的发现期和657例和1183例对照的复制期的关联结果(P<0.05)
|
|
|
发现 |
重复 |
||||
|
类别 |
HLA等位基因 |
频率_ |
或(95% 置信区间) |
P |
频率b |
OR |
P |
|
I |
A*0201 |
0.26 |
1.35 (1.05, 1.74) |
0.020 |
0.28 |
1.07(0.89, 1.28) |
0.48 |
|
|
A*0301 |
0.15 |
0.66 (0.46, 0.94) |
0.023 |
0.14 |
1.00(0.79, 1.27) |
1.0 |
|
|
B*0801 |
0.11 |
0.60(0.38, 0.92) |
0.021 |
0.10 |
0.75(0.52, 1.08) |
0.12 |
|
|
|||||||
|
II |
DRB1*0301 |
0.12 |
0.52 (0.34, 0.81) |
3.2×10 -3 |
0.12 |
0.67 (0.50, 0.91) |
0.011 |
|
|
DQA1*0501 |
0.25 |
0.74 (0.56, 0.97) |
0.031 |
0.28 |
0.81 (0.67, 0.99) |
0.037 |
|
|
DQB1*0201 |
0.12 |
0.51 (0.33, 0.79) |
2.7×10 -3 |
0.12 |
0.70 (0.52, 0.94) |
0.018 |
a.发现数据集控件中 的等位基因频率
b复制数据集控制中的等位基因频率
在 657 个非重叠骨肉瘤病例和 1183 个欧洲血统对照的复制数据集中,MHC 区域中的总共 5,401 个 SNP 在阵列上成功进行基因分型并用于估算四位数的 HLA 变体。发现集中鉴定的 HLA I 类变体均未与复制数据中的骨肉瘤风险相关(P范围= 0.12-1.0)。然而,发现数据集中鉴定的所有三个 HLA II 类变体都显示了复制数据中关联的证据(P范围= 0.011-0.037)(表1)。
两项研究的荟萃分析确定 HLA-DRB1*0301 为最高信号(OR meta =0.62;95% CI = 0.48–0.79;P meta =1.5×10 -4),达到 Bonferroni 校正显着性。发现分析中发现的其他 II 类变体(即DQA1*0501 和 DQB1*0201)具有相似的 p 值和效应大小(表 2)。骨肉瘤风险基因检测方案开发小组观察到这三个 II 类基因(DQB1*0201、DRB1*0301 和 DQA1*0501)的受试者基因型高度相关(r 2 =0.33-0.98)。在以 DRB1*0301 基因型为条件的逻辑回归分析中,在荟萃分析中,DQA1*0501 和 DQB1*0201 均与骨肉瘤风险显着相关(P范围=0.35-0.97),表明这三个等位基因代表单一保护性单倍型.
表 2:名义上与骨肉瘤相关的 HLA II 类变体的 Meta 分析结果 (p<0.05),在 3 个统计学上独立的位点
|
HLA等位基因 |
频率发现_ |
ORmeta(95% CI) |
Pmetab |
I 2c (%) |
信号 |
|
DRB1*0301 |
0.12 |
0.62 (0.48, 0.79) |
1.5×10 -4 |
0 |
1 |
|
DQA1*0501 |
0.25 |
0.79 (0.67, 0.92) |
3.2×10 -3 |
0 |
1 |
|
DQB1*0201 |
0.12 |
0.63 (0.49, 0.81) |
2.6×10 -4 |
23.8 |
1 |
|
DRB1*0101 |
0.08 |
1.37 (1.10, 1.71) |
4.9×10 -3 |
0 |
2 |
|
DQA1*0101 |
0.15 |
1.33 (1.12, 1.58) |
1.2×10 -3 |
0 |
2 |
|
DQB1*0501 |
0.13 |
1.34 (1.11, 1.61) |
2.5×10 -3 |
0 |
2 |
|
DQB1*0302 |
0.11 |
0.73 (0.58, 0.91) |
6.4×10 -3 |
0 |
3 |
a发现数据集控件中的等位基因频率,在复制集中具有相似的频率
b针对祖先成分调整的固定效应荟萃分析 p 值。粗体突出显示 Bonferroni 校正后的显着性,并调整了变体之间的连锁不平衡 ( P <1.3×10 -3 )
c I 2异质性指数
荟萃分析还揭示了另外三个 II 类等位基因的另一个关联信号:DRB1*0101 (OR meta =1.37;95% CI=1.10–1.71;P meta =4.9×10 -3 ),DQA1*0101 (OR meta = 1.33;95% CI=1.12–1.58;P meta =1.2×10 -3)和DQB1*0501(OR meta =1.34;95% CI=1.11–1.61;P meta =2.5×10 -3)。这三个等位基因之间的高 LD (r 2= 0.57–0.99),以及显示信号衰减的条件分析表明它们代表单一风险单倍型。调整位于上述保护性单倍型上的 HLA-DRB1*0301 的条件分析并未显着减弱与这三个等位基因的关联(P范围= 6.3×10 -3 - 0.014),表明它们代表第二个独立信号。
骨肉瘤风险基因检测方案开发小组在 DQB1*0302 观察到第三个独立风险位点,提示与骨肉瘤相关(OR meta =0.73;95% CI=0.58-0.91;P meta =6.4×10 -3)。调整 DRB1*0301(保护单倍型中的最高信号)和 DQA1*0101(风险单倍型中的最高信号)的条件分析并未显着减弱 DQB1*0302 的关联(P条件= 0.01). 此外,当调整 rs1906953 时,所有三个 HLA II 类关联信号仍然存在,6p21.3 上的GRM4变体先前被确定为骨肉瘤风险基因座,这表明骨肉瘤风险基因检测方案开发小组的任何 HLA II 类风险变体都没有 LD (r 2 <0.01)。
尽管复制数据集基因分型阵列不包含发现数据集自定义阵列中包含的 MHC 区域的额外约 6,000 个探针,但对于上面报告的名义上显着骨肉瘤风险基因座的发现和复制数据集,插补精度同样高。
对组合发现和复制数据的性别特异性分析表明,第三个 HLA II 类信号 DQB1*0302 的影响可能会因受试者性别而改变(P交互作用=0.02)。具体而言,骨肉瘤风险基因检测方案开发小组观察到女性(n=397;OR meta =0.54;P meta =2.4×10 -3)比男性(n=506;OR meta =0.90;P meta=0.49)。对于其他关联信号,未观察到受试者性别不同层次的差异。在可获得临床数据的发现样本的仅病例分析中,顶部信号的先导 HLA 变体 (DQB1*0201) 与临床特征没有显着相关性,尽管它暗示与更高的肿瘤分级相关(P趋势= 0.09 )。
对该地区非加性效应的系统搜索确定了 HLA-DQB1*0302 处的可加性偏离。包含一个优势项提高了模型在发现和复制数据集中的拟合度(分别为 P改进= 0.04 和 P改进= 0.03)。显性项(OR=0.48;95% CI=0.25–0.92)表明,对于 DQB1*0302,杂合性提供的保护作用是预期的两倍,在该等位基因纯合的个体中观察到的效果。鉴于 DQB1*0302 具有女性特异性关联,骨肉瘤风险基因检测方案开发小组进行了性别分层的非加性分析,观察男性受试者的非加性保护作用(P改善=0.09 和 P改善=6.4×10 -3对于发现和复制数据集),但不是女性受试者(对于发现和复制数据集,P提高=0.23,P提高=0.51)。对于任何其他 DQB1 等位基因或任何其他 HLA II 类等位基因,均未观察到非加性效应。以 DRB1*0301 为代表的顶部信号的上位性分析确定了与连锁等位基因 DRB1*1301(P相互作用=3.0×10 -3)、DQA1*0103(P相互作用=2.5×10 -3)和 DQB1*的暗示相互作用0603 (P交互作用=4.8×10 -3 ),尽管在调整所有测试的交互作用后这些交互作用不显着 (即0.05/26=1.9×10 -3)。没有证据表明其他顶级基因座出现相互作用表 2被观测到。
先前关于 HLA 变体与肠易激疾病之间关联的研究表明,肽结合沟处的静电电位差异与 DRB1 等位基因对疾病风险的影响方向性相关 。骨肉瘤风险基因检测方案开发小组查询了他们公布的 DRB1*0301 和 DRB1*0101 的静电势数据,骨肉瘤风险基因检测方案开发小组的两个顶部信号显示出相反的效果。骨肉瘤风险基因检测方案开发小组观察到这两个等位基因之间的静电势差约为 0.8,范围从 0(相同)到 1(最大差异),表明骨肉瘤风险等位基因 (DRB1*0101) 和保护性等位基因 (DRB1*0301)不同的静电势,支持这些等位基因之间的潜在功能差异。
在估算的HLA基因内SNP的二次分析中,骨肉瘤风险基因检测方案开发小组观察到HLA-A下游的几个SNP显示出与骨肉瘤风险相关的证据,包括先导SNP rs2517612(OR meta =0.60,P meta =5.5×10 -5)。在 HLA-DRB1 上游的 rs3129890 处观察到一个额外的独立信号(OR meta =0.71,P meta =5.7×10 -5)。基因内 SNP 处于中度 LD,具有最高的 HLA II 类信号 DRB1*301 (r 2 =0.41-0.42)。然而,对于多次测试(即<2.9×10 -5),两个SNP都没有幸存下来。
骨肉瘤风险基因的分析
在迄今为止的第一项关于 HLA 变异与骨肉瘤风险之间关联的研究中,骨肉瘤风险基因检测方案开发小组观察到一个与风险降低相关的信号,该信号由三个相关的等位基因 DQB1*0201、DRB1*0301 和 DQA1*0501 组成,在独立数据集中进行了复制在 Bonferroni 修正后仍然显着。发现和复制数据集的荟萃分析揭示了另外两个独立信号,一个是 DQB1*0302 风险降低,另一个是三个相关等位基因 DRB1*0101、DQA1*0101 和 DQB1*0501 风险增加。在调整 rs1906953 的条件分析中,这三个信号中没有一个被减弱,表明这些影响与GRM4中 6p21.3 处先前确定的 GWAS 信号无关。
关于骨骼和免疫细胞之间复杂相互作用的文献越来越多,它们具有共享的信号转导途径、共同的造血前体以及骨骼和免疫系统之间的交互作用 。多项研究已观察到 HLA II 类分子在人类成骨细胞上的表达 ,正如骨肉瘤中 HLA II 类受体活性通路的上调 ,支持 HLA II 类基因在骨肉瘤中的生物学作用发展和进步。事实上,除了目前正在临床开发的骨肉瘤患者的免疫治疗靶点(例如PD1、PDL1)、HLA II 类分子、IL-10 和 TGFβ 目前被认为是破坏对肿瘤的免疫耐受的潜在靶标 。
最近的一项研究发现,免疫细胞转录本的变异,特别是单核细胞和 T 细胞的变异与骨肉瘤的转移和存活之间存在关联 。免疫细胞浸润和免疫监测在骨中的作用与骨肉瘤风险基因检测方案开发小组的结果一致,表明对骨肉瘤病因的潜在免疫学贡献,可能通过抗原呈递的差异起作用。尽管骨肉瘤风险基因检测方案开发小组无法评估患者的存活率,但 DQB1*0201 与肿瘤分级之间的暗示性关联表明免疫相关基因的种系变异可能会影响患者的预后。这补充了最近的 GWAS,该 GWAS 确定了与 IL-33 表达降低和骨肉瘤患者存活率降低相关的等位基因。IL-33 诱导 Th2 相关细胞因子的产生,并与特应性疾病有关 。因为它由成骨细胞和淋巴细胞表达,局部和全身免疫变化在影响骨肉瘤风险和结果方面的相互作用以及 IL-33 和 HLA II 类变异之间的相互作用值得进一步关注。
对其他肿瘤部位的研究也揭示了骨肉瘤风险基因检测方案开发小组的结果中突出显示的 HLA 等位基因的参与,提出了这种变异作为遗传多效性来源的有趣可能性。值得注意的是,与骨肉瘤风险基因检测方案开发小组的最高关联信号相对应的扩展单倍型 DRB1*0301:DQA1*0501:DQB1*0201 先前与卵巢癌风险增加有关 。发现与 Epstein-Barr 病毒相关的鼻咽癌风险增加与扩展单倍型 HLA-A*3303-B*5801/2-DRB1*0301-DQB1*0201/2-DPB1*0401,特定于中国人台湾研究族群。先前在 B 细胞恶性肿瘤中描述了 MHC 区域内多效性关联的例子,这表明共享生物学途径可能影响不同癌症类型的发展 。此外,之前已经注意到不同 B 细胞恶性肿瘤中的相反风险关联与 MHC 区域中的多效性风险基因座 相似,类似于骨肉瘤风险基因检测方案开发小组观察到与相同 HLA 变体的其他癌症类型风险增加相比对骨肉瘤的保护作用。骨肉瘤风险基因检测方案开发小组第二个信号的单个等位基因 DRB1*0101、DQA1*0101、DQB1*0501 也与多种癌症有关,包括肾细胞癌、乳腺癌、宫颈癌和儿童急性淋巴细胞白血病。骨肉瘤风险基因检测方案开发小组的第三个信号 DQB1*0302 与 HPV 相关的宫颈癌有关,推测是由于对 HPV 抗原结合的不同亲和力与保护免受感染有关 。
对 HLA 基因内 SNP 的探索性分析确定了先导 SNP rs2517612,这是大量基因的表达数量性状基因座 (eQTL),包括:BTN3A2、SLC15A3、FAM20B、HLA-B、HLA-C、HLA-DQA1、HLA-DQA2、其中。由于分析了大量的 HLA 基因内 SNP,没有单个 SNP 关联在 Bonferroni 校正中幸存下来。
DQB1*0302 与受试者性别之间的潜在相互作用是有趣的,因为有充分证据表明男性与女性的骨肉瘤发病率增加。尽管样本量较小,但在具有 DQB1*0302 等位基因的女性中,与男性相比,骨肉瘤风险的更大和更显着降低与这种可能导致女孩骨肉瘤发病率降低的变异一致。尚不清楚为什么这种关联在女性中会更强,尽管之前曾观察到胃癌和急性淋巴细胞白血病的性别特异性 HLA 关联。一种可能性可能与青春期骨骼生长速率的性别差异有关,以前认为由于快速增殖的细胞对致癌剂和有丝分裂错误的敏感性,会影响骨肉瘤的风险。然而,观察到的男性特异性非加性保护作用使解释复杂化,并表明 DRB1*0302 在两性骨肉瘤易感性中的作用。由于这种 HLA 变异在 Bonferroni 校正后并未保持显着性,因此需要进行额外的后续研究以确认关联并探索性别的任何影响修饰。
DQB1*0302 关联在发现和复制数据集中都显示出非加性优势效应,这意味着杂合子中的保护作用比纯合子更强。假设 MHC 区域的非加性效应在免疫系统必要的功能冗余中发挥重要作用,并且考虑到骨骼和免疫系统之间的重叠信号转导途径和串扰,检测这种与骨肉瘤有关的影响很有趣。
因为骨肉瘤风险基因检测方案开发小组的基因分型数据是使用定制的 SNP 阵列生成的,而不是通过靶向测序直接分型 HLA 等位基因,所以骨肉瘤风险基因检测方案开发小组的基因分型受到插补正确性的限制。然而,插补错误只会导致检测信号的能力降低,而不会导致 I 类错误率的任何增加。除了罕见的变异,HLA 插补也可能无法检测旁系同源基因和拷贝数可变区中的变异,因为这些变异可能由于偏离 Hardy-Weinberg 平衡而被排除在分析之外。更高分辨率的 HLA 测序对于更明确地识别骨肉瘤风险基因检测方案开发小组观察到的关联背后的致病变异是必要的,这可能使特定的保护性 HLA 变异能够指导打破对肿瘤免疫耐受的方法。
使用的基于阵列的平台也限制了骨肉瘤风险基因检测方案开发小组检测与遗传性癌症易感综合征相关的基因中罕见或私人生殖系突变的能力,包括TP53。虽然之前的研究没有观察到常见的TP53变体与骨肉瘤风险之间存在很强的关联性 ,但最近的一项研究发现,在 30 岁之前诊断出的骨肉瘤患者中,有 9.5% 至少存在一种罕见的外显子TP53变体 。鉴于最近有报道称,未选择癌症病史的对照组中 TP53 突变的人群患病率高于预期,以及多种罕见变异对肉瘤风险的潜在多基因贡献,应探索 MHC II 类变体和其他免疫因子在改变TP53突变携带者癌症外显率中的作用。
骨肉瘤风险基因检测方案开发小组的研究有几个额外的限制。骨肉瘤风险基因检测方案开发小组的分析是在欧洲血统的受试者中进行的,结果可能无法推广到其他种族。SNP2HLA 方法需要特定种族的参考面板,目前缺乏足够样本量的非欧洲参考数据集。尽管骨肉瘤风险基因检测方案开发小组对 DRB1 变体的静电特性的查询表明,这两个信号的 DRB1 等位基因是功能相关性的强有力候选者,但骨肉瘤风险基因检测方案开发小组也不能自信地确定在骨肉瘤风险基因检测方案开发小组的前两个信号中的多个连锁 HLA 变体中最可能的致病等位基因。然而,需要额外的功能研究来推断因果关系。最后,虽然骨肉瘤风险基因检测方案开发小组的两个主要信号在 LD 调整后的 Bonferroni 校正多次测试后仍然显着,但需要额外的数据集来确认骨肉瘤风险基因检测方案开发小组的发现。
总之,骨肉瘤风险基因检测方案开发小组以 4 位分辨率对 HLA I 类和 II 类等位基因进行了基因分型,并观察到多个与儿童骨肉瘤风险相关的独立 II 类等位基因,包括 DRB1 的两个不同等位基因。这些发现提高了骨肉瘤风险基因检测方案开发小组对骨肉瘤病因的潜在免疫学贡献的理解,也可能为未来免疫疗法的发展提供见解。
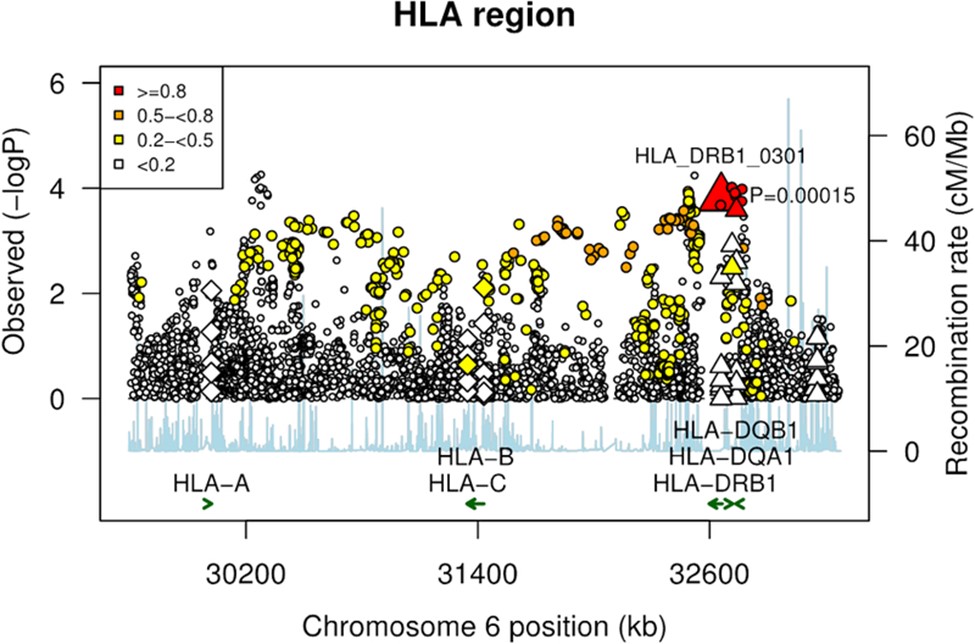
图1:HLA 变体荟萃分析的区域关联图。
菱形代表 HLA I 类变体,三角形代表 HLA II 类变体,圆圈代表 HLA 基因内 SNP。大三角形代表 DRB1*0301 中的顶部信号。
Two HLA Class II Gene Variants Are Independently Associated with Pediatric Osteosarcoma Risk.
Zhang C, Wiemels JL, Hansen HM, Gonzalez-Maya J, Endicott AA, de Smith AJ, Smirnov IV, Witte JS, Morimoto LM, Metayer C, Walsh KM.
Cancer Epidemiol Biomarkers Prev. 2018 Oct;27:1151-1158. doi: 10.1158/1055-9965.EPI-18-0306. Epub 2018 Jul 23.
- 【佳学基因检测】唇裂、腭裂的基因检查与检测分析...
- 【佳学基因检测】怎么筛查唇腭裂?...
- 【佳学基因检测】唇裂与腭裂基因检测...
- 【佳学基因检测】基因检测说明唇腭裂是否是因为怀孕期间吃了什么东西?...
- 【佳学基因检测】唇腭裂基因检测可以找到畸形发生的原因吗?...
- 【佳学基因检测】常见骨科基因检测项目,这些疾病会遗传!...
- 【佳学基因检测】哪些骨头病会遗传,如何做基因检测才能阻断遗传,帮助治疗?...
- 【佳学基因检测】骨科遗传性疾病基因检测...
- 【佳学基因检测】什么时候会诊断为疑似戈谢病?怎么做基因检测分析?...
- 【佳学基因检测】软组织和骨肿瘤基因检测分子病理学共识与标准...
- 【佳学基因检测】骨科疾病为什么要做染色体基因检测?检测结果如何指导遗传阻断?...
- 【佳学基因检测】用病例说明早衰症基因检测是怎么做的?...
- 【佳学基因检测】骨穿基因检测是什么?...
- 【佳学基因检测】 骨科检查项目及费用...
- 【佳学基因检测】骨科基因检测项目有哪些?...
- 【佳学基因检测】基因检测如何找到骨关节炎效果基因?...
- 【佳学基因检测】脊柱侧弯基因检测细分型、好治疗!...
- 【佳学基因检测】并指症基因检测,怎样才能知道如何阻断遗传?...
- 【佳学基因检测】多了一个指头是病吗,是否遗传,基因检测给答案!...
- 【佳学基因检测】遗传性佝偻病基因检测,优质机构推荐...
- 【佳学基因检测】遗传性矮小症基因检测...
- 【佳学基因检测】骨代谢性疾病基因检测...
- 【佳学基因检测】先天性软骨病基因检测及其对治疗的帮助作用...
- 【佳学基因检测】揭秘“瓷娃娃”成骨不全症,基因检测如何给予帮助?...
- 【佳学基因检测】成骨不全症基因检测(Osteogenesis imperfecta)...
- 【佳学基因检测】骨质疏松需要做基因检测吗?...
- 【佳学基因检测】抗骨质疏松药用药安全基因检测:怎么做?...
- 【佳学基因检测】大理石骨病基因检测如何帮助治疗...
- 【佳学基因检测】骨石症基因检测如何选择可信赖的机构...
- 【佳学基因检测】硬骨症基因检测及其治疗方案...
- 来了,就说两句!
-
- 最新评论 进入详细评论页>>
